The Plasma Membrane: Structure
Plasma Membrane
The flexible plasma membrane defines the barrier of a cell by separating two of the body’s major fluid compartments – the intracellular fluid within cells and the extracellular fluid (ECF) outside cells. The term cell membrane is commonly used as a synonym for plasma membrane, but in this article we will refer to the cell’s surface as the plasma membrane. The plasma membranes unique permeable structure allows it to play a dynamic role in cellular activities.
The Fluid Mosaic Model
The fluid mosaic model of membrane structure depicts the plasma membrane as an exceedingly thin structure composed of a double layer, or bilayer, of lipid molecules with protein molecules dispersed within its layer. The proteins, many of which float in the fluid lipid bilayer, form a constantly changing mosaic pattern. The model is named for this characteristic.
Membrane Lipids
The lipid bilayer forms the basic “fabric” of the membrane. It is constructed largely of phospholipids, with smaller amounts of glycolipids, cholesterol, and areas called lipid rafts.
Phospholipids
Each phospholipid molecule has a polar “head” that is charged and is hydrophilic (water-loving), and an uncharged, nonpolar “tail” that is made of two fatty acid chains and is hydrophobic (water-fear). The polar heads are attracted to water – the main constituent of both the intracellular and extracellular fluids – and so they lie on both the inner and outer surfaces of the membrane. The nonpolar tails, being hydrophobic, avoid water and line up in the center of the membrane.
The result is that the plasma membranes, indeed all biological membranes, share a sandwich-like structure: They are composed of two parallel sheets of phospholipid molecules lying tail to tail, with their polar heads exposed to water on either side of the membrane or organelle. This self-orienting property of phospholipids encourages biological membranes to self-assemble into closed, generally spherical, structures and to reseal themselves when torn.
With a consistency similar to olive oil, the plasma membrane is a dynamic fluid structure in constant flux. Its lipid molecules move freely from side to side, parallel to the membrane surface, but their polar-nonpolar interactions prevent them from flip-flopping or moving from one half of the bilayer to the other half. The inward-facing and outward-facing surfaces of the plasma membrane differ in the kinds and amounts of lipids they contain, and these variations are important in determining local membrane structure and function. Most membrane phospholipids are unsaturated, a condition which kinks their tails and increases membrane fluidity.

The lipid bilayer forms the basic structure of the membrane. The associated proteins are involved in membrane functions such as membrane transport, catalysis, and cell-to-cell recognition.
Glycolipids
Glycolipids are lipids with attached sugar groups. Found only on the outer plasma membrane surface, glycolipids account for about 5% of total membrane lipids. Their sugar groups, like the phosphate-containing groups of phospholipids, make the end of the glycolipid molecule polar, whereas the fatty acid tails are nonpolar.
Cholesterol
Some 20% of membrane lipid is cholesterol. Like phospholipids, cholesterol has a polar region and a nonpolar region. It wedges its platelike hydrocarbon rings between the phospholipid tails, stabilizing the membrane, while decreasing the mobility of the phospholipids and the fluidity of the membrane.
Membrane Proteins
A cell’s plasma membrane bristles with proteins that allow it to communicate with its environment. Proteins make up about half of the plasma membrane by mass and are responsible for most of the specialized membrane functions. Some membrane proteins float freely. Others are “tethered” to intracellular structures that make up the cytoskeleton and are restricted in their movement.
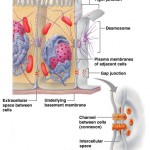
There are two distinct populations of membrane proteins, integral and peripheral.
Integral Proteins
Integral proteins are firmly inserted into the lipid bilayer. Some protrude from one membrane face only, but most are transmembrane proteins that span the entire membrane and protrude on both sides. Whether transmembrane or not, all integral proteins have both hydrophobic and hydrophilic regions. This structural feature allows them to interact with both the nonpolar lipid tails buried in the membrane and the water inside and outside the cell.
Some transmembrane proteins are involved in transport, and cluster together to form channels, or pores, through which small, water-soluble molecules or ions can move, thus bypassing the lipid part of the membrane. Others act as carriers that bind to a substance and then move it through the membrane. Some transmembrane proteins are enzymes. Still others are receptors for hormones or other chemical messengers and relay messages to the cell interior – a process called signal transduction.
Peripheral Proteins
Unlike integral proteins, peripheral proteins are not embedded in the lipid bilayer. Instead, they are attached loosely to integral proteins and are easily removed without disrupting the membrane. Peripheral proteins include a network of filaments that help support the membrane from its cytoplasmic side. Some peripheral proteins are enzymes. Others are motor proteins involved in mechanical functions, such as changing cell shape during cell division and muscle cell contraction. Others link cells together.
Lipid Rafts
About 20% of the outer membrane surface contains lipid rafts, dynamic assemblies of saturated phospholipids associated with unique lipids called sphingolipids and lots of cholesterol. The quiltlike lipid rafts are more stable and less fluid than the rest of the membrane, and they include or exclude specific proteins to various extents. Because of these qualities, lipid rafts are assumed to be concentrating platforms for certain receptor molecules or for protein molecules needed for cell signaling and other functions.
The Glycocalyx
Many of the proteins that abut the extracellular fluid are glycoproteins with branching sugar groups. The term glycocalyx describes the fuzzy, sticky, carbohydrate-rich area at the cell surface. Think of your cell as sugar coated. The glycocalyx on each cell’s surface is enriched both by glycolipids and by glycoproteins secreted by the cell.
Because every cell type has a different pattern of sugars in its glycocalyx, the glycocalyx provides highly specific biological markers by which approaching cells recognize each other. For example, a sperm recognizes an ovum by the ovum’s unique glycocalyx. Cells of the immune system identify a bacterium by binding to certain membrane glycoproteins in the bacterial glycocalyx.
Check your understanding
- What basic structure do all cellular membranes share?
- Why do phospholipids, which form the greater part of membranes, organize into a bilayer – tail-to-tail – in a watery environment?
- What is the importance of the glycocalyx in cell interactions?
Category: Cells




